El ojo
Clasificado en Otras materias
Escrito el en  español con un tamaño de 27,52 KB
español con un tamaño de 27,52 KB
1. Estrutura y ultraestructura de los conos y bastones.
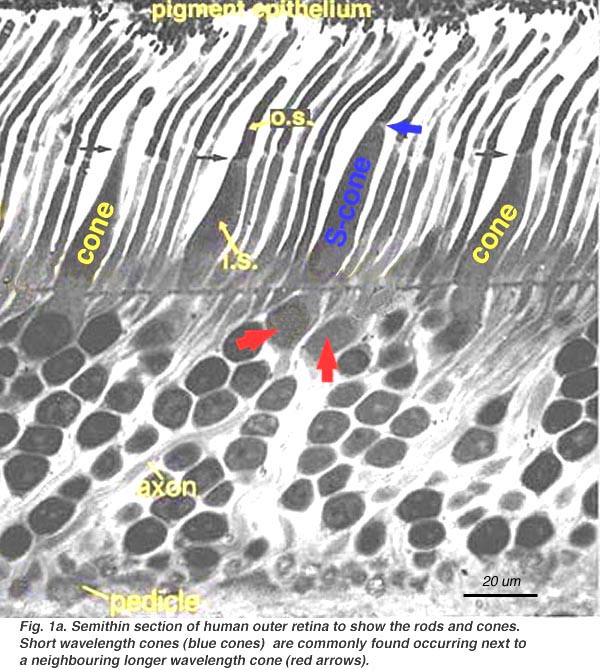
En una sección vertical de una retina vista al microscopio óptico es relativamente fácil distinguir a los conos de los bastones.
Los conos presentan una estructura cónica, con sus núcleo alineados en una sóla capa justo por debajo de la membrana limitante externa. Sus segmentos internos y externos se proyectan dentro del espacio subretinal hacia el epitelio pigmentario. A nivel de la fovea, donde sólo existen conos, sus cuerpos celulares se situan oblicuamente con respecto a sus procesos. Los bastones por otra parte, poseen una morfología alargada con sus segmentos internos y externos rellenado el espacion entre los conos y los procesos de las células del epitelio pigmentario. Los cuerpos celulares de los bastones constituyen el resto de la capa nuclear externa, donde se situan formando varias capas. Aunque no siempre resulta claro en las preparaciones histológicas habituales, los procesos de las células del epitelio pigmentario rodean completamente tanto a los segmentos externos de los conos como los de los bastones.
Utilizando el microscopio electrónico se obtiene una mejor resolución de la morfología de los conos y bastones. Es obvio que los segmentos internos (i.s.) de los bastones son más delgados que los de los conos. Así los segmentos internos de los conos poseen un diámetro de unas 6 micras frente a las 2 micras de los segmentos internos de los bastones. Sin embargo a nivel de la fovea, donde sólo existe conos, los segmentos internos de estos pueden llegar a medir unicamente alrededor de 1.5 micras. Los segmentos internos de los conos y bastones estan repletos de largas mitocondrias. A nivel de la unión entre los segmentos internos y externos de los fotoreceptores existe un cilio de unión.
A partir de estos cilios se producen una serie de evaginaciones e invaginaciones de la membrana plasmática de los fotoreceptores que dan lugar a los segmentos externos (o.s.). Esta es la porción de los fotoreceptores donde se encuentran los pigmentos visuales.Los segmentos externos de los conos y bastones derivan de repliegues de la membrana plasmática (ver Fig. 4,5 y la animación que les sigue). .
Al final a nivel de los bastones, los segmentos externos estan constituidos por discos membranos aislados de la membrana plasmática donde se encuentran inmersos los pigmentos sensibles a las radiaciones luminosas. Por contra en los conos no existen discos membranosos aislados sino multiples repliegues de la membrana plasmática.
La rodopsina es el pigmento visual que se encuentra nivel de los segmentos externos de los bastones. Esta formada por una molecula proteica, la opsina, que se fabrica en el aparato de Golgi (situado en los segmentos internos) y el retinal. La opsina se dirige hacia la zona del cilio de unión gracias a la acción de proteinas G y desde aquí pasa ya hacia el segmento externo (Papermaster et al., 1985; Derectic and Papermaster, 1995).La otra parte del pigmento visual, el retina (derivado de la vitamina A) en proporcionado a los discos desde el epitelio pigmentaria a través de proteinas transportadoras ( proteínas IRPB) que se encuentran a nivel de la matriz que existe entre los distintos fotoreceptores.
2. Pigmentos visuales y fototransducción
Los fotoreceptores de todos los vertebrados responden a la luz en función de los pigmentos visuales que se encuentran incrustados a nivel de la bicapa lipídica de los repliegues (en el caso de los conos) y discos membranosos (en el caso de los bastones).
Los bastones contienen rodopsina y son responsable de la visión en condiciones de baja luminosidad, presentando un pico de mayor sensibilidad hacia la longitud de onda de los 500 nm (luz verde azulada). Los conos por su parte contienen tres tipos diferentes de opsinas. Una con mayor sensibilidad para las longitudes de onda largas (luz roja), otra que es sensible a las longitudes de onda medias (luz verde) y otra con mayor sensibilidad a las longitudes de onda cortas (luz azul). Los conos son la base de la percepción del color.
Cada molécula de rodopsina consiste en siete porciones transmembranosas que rodean al 11-cis retinal. Este 11-cis retinal o cromóforo se une mediante un residuo de lisina a la septima helice (Hargrave et al., 1984; Hargrave and McDowell, 1992). Cada disco de los segmentos externos continen miles de estas moléculas. Cuando un foto de luz llega a esta nivel el cromoforo se isomeriza y pasa de la forma 11-cis a la forma todo trans, lo cual da lugar a cambios conformacionales de la proteina, que producen lo que se denomina como blanqueamiento de la rodopsina. Durante este proceso se forman varios metabolitos intermediarios como la Metarodopsina II que activa a una proteina G especial, conocida como Transducina que al final va a desencadenar la cascada de la fototransducción que se presenta en la Fig. 10.
Cuando la retina esta en condiciones de oscuridad, se encuentran abiertos una serie de canales ionicos a nivel de los segmentos externos de los fotoreceptores que permiten la entrada fundamentalmente de iones Sodio. Esta entrada de Sodio, depolariza parcialmente a los fotoreceptores, permitiendo la liberación de neurotransmisor a nivel de sus terminales sinápticos. El transmisor liberado se supone que es Glutamato. Cuando la luz estimula a la molecula de rodopsina, se producen una sería de cambios que se presentan esquemáticamente en la Fig. 9. y en la animación que la sigue, que van a producir el cierre de los canales ionicos permeables al Sodio. Por tanto cesa la entrada de Sodio y el fotoreceptor se hiperpolariza, con lo que deja de liberar neurotransmisor (para profundizar en los mecanismos de este proceso consultar las revisiones de Stryer, 1991; Yau, 1994 y Kawamura, 1995).
La corriente que se produce durante las condiciones de oscuridad es debida en un 80% a la entrada de iones Sodio, sin embargo el canal también es permeable para los iones de Calcio y Magnesio (Yau, 1994). Además en oscuridad debe existir un mecanismo para eliminar tanto el Calcio como el exceso de Sodio. Este mecanismo parece ser que consiste en un intercambiador Sodio/Calcio a nivel de la membrana de los segmentos externo. El Calcio, además tiene un importante papel en todo el proceso de la fototransducción, ya que aunque no participa directamente en la cascada de la fototransducción, mejora la capacidad de los bastones para recuperarse después de la iluminación, teniendo un importante papel regulador en los fenomenos de adaptación a las condiciones de luz/oscuridad (Yau, 1994).3. Fagocitosis de los segmentos externos por las células del epitelio pigmentario.
Los discos membranosos que contienen los pigmentos visuales estan continuamente renovandose en el caso de los bastones. Nuevos discos son añadidos a nivel de la unión de los segmentos interno y externo que van desplazando hacia la zona del epitelio pigmentario a los discos viejos. Estos discos más externos son fagocitados por las células del epitelio pigmentario durante el ciclo diurno y convertidos en fagosomas.
En el caso de los conos las células del epitelio pigmentario también fagocitan sus porciones más externas durante el ciclo diurno, pero en diferentes periodos del día. Asi en el caso de los bastones la fagocitosis se produce fundamentalmente hacia la hora de la salida del sol, mientras que en el caso de los conos los procesos de fagocitosis aumentan cuando se acerca la hora de la puesta de sol (Young, 1971, 1976; Le Vail, 1976; Steinberg et al., 1977; Beharse, 1982).
4. Tipos de conosA diferencia de los bastones, que forman un sólo tipo morfológico y funcional de fotoreceptor, existen tres tipos de conos: unos que presentan una sensibilidad máxima para las longitudes onda más largas ("conos rojos"), otros con mayor sensibilidad a las longitudes de onda medias ("conos verdes") y otros con mayor sensibilidad a las longitudes de onda más cortas ("conos azules"). Estos tres tipos de conos dan lugar a la visión tricomática que poseen la mayoría de los humanos.
Estudios fotométricos y psicofisiológicos han demostrado que en la retina humana los conos rojos tienen su pico de sensibilidad a los 558 nm, los conos verdes a los 531 nm y los conos azules a los 420 nm (consultar el trabajo de Gouras, 1984 para una revisión más amplia).A diferencia de los seres humanos, algunas especies de mamiferos poseen una visión dicromática debido a la presencia de bastones y de sólo dos tipos de conos: los sensibles a las longitudes de onda medias y cortas. Por el contrario otros animales como las aves, reptiles y peces poseen mecanismos de visión tricomátrica e incluso pentacromática
Aunque en retina de aves, peces y reptiles existen algunas diferencias morfológicas entre los diversos tipos de conos, no esta claro que ocurra lo mismo en la retina de los primates. Sin embargo parece ser que si existen evidencias que muestran que es posible distinguir al menos a los conos azules del resto de los conos utilizando métodos exclusivamente morfológicos (Ahnelt et al., 1987).
5. Densidad de conos y bastones en retina humana.
Para entender la organización de los circuitos neuronales dentro de la retina es preciso conocer la organización espacial de los distintos tipos de fotoreceptores a lo largo de la retina. Las Figuras 15 y 16 muestran las densidades para cada uno de los tipos de fotoreceptores. Así en la fovea existe una alta densidad de conos que se encuentran distribuidos espacialmente formando un mosaico hexagonal muy regular. Por fuera de la fovea, la presencia de bastones desorganiza un poco este patrón hexagonal. En terminos cuantitativos, la mayor densidad de conos se concentra a nivel de la foveola, decreciendo su numero conforme nos alejamos de la misma hasta una densidad mas o menos uniforme en la retina periférica (Osterberg, 1935; Curcio et al., 1987). Existe también un pico de bastones alrededor de la fovea. La zona de la papila (que corresponde al nervio óptico) carece de cualquier tipo de fotoreceptor ("punto ciego").6. Ultraestructura de las terminaciones sinápticas de los conos y los bastones.
La información que codifican los fotoreceptores, respecto al número de quanta de luz y su sensibilidad respecto a las distintas longitudes de onda es transmitida a través de sus terminaciones sinápticas. Estas terminaciones son de forma triangular y se denominan pediculos en el caso de los conos, mientras que su morfología es redondeada, denominandose esférulas en el caso de los bastones.
Ambos tipos de terminaciones estan rellenas de vesículas sinápticas. Además a nivel de las sinápsis con los siguientes tipos celulares (células horizontales y bipolares) contienen unas estructuras densas que se conocen como Sinapsis en Cintilla. Cada pediculo presenta aproximadamente unas 30 de estas sinápsis en cintilla (Ahnelt et al., 1990) mientras que las esférulas contienen sólo unas 2 de estas sinápsis en cintilla.A nivel de los pediculos se forman una estructuras que se conocen como triadas en la que encontramos 3 procesos: 2 procesos laterales que corresponden a células horizontales (HC) y un proceso central, alineado con la sinápsis en cintilla que corresponde a una célula bipolar invaginante (imb). Además existen otros tipos de células bipolares que también contactan con el pediculo mediante contactos basales (fmb) (Missoten, 1965; Dowling y Boycott, 1966; Kolb, 1970).
Las esférulas de los bastones presentan sólo 2 sinápsis en cintilla, que dan lugar a unas estructuras conocidas como diadas en las que existen 1 elemento lateral y un elemento central. Los elementos laterales son terminaciones axonicas de las celulas horizontales (HC)mientras que los elementos centrales corresponden a dendritas invaginantes de las células bipolares para bastones (Missoten, 1965; Dowling y Boycott, 1966; Kolb, 1970). No existen habitualmente contactos basales a nivel de las esferulas de los bastones.
Además de estos tipos de contactos sinápticos también existen sinápsis de tipo electrico entre conos y conos y entre conos y bastones a nivel de la retina humana. Así los pediculos de los conos presentan una pequeñas proyecciones laterales que se denominan telodendria y que forman pequeñas sinapsis eléctricas (Raviola and Gilula, 1975; Nelson et al., 1985).
La esclerótica es la cápsula fibrosa más externa del globo ocular, que por su parte anterior se vuelve transparente y da lugar a la córnea.
Pero la esclerótica, no es exclusivamente blanca, sino que presenta numerosos capilares, importantes en el diagnóstico de problemas de tipo circulatorio.
Según la mayoría de autores, la presencia de un vaso que acaba cerca de un sector determinado del iris, o bien de un vaso en forma de V, o con un codo derca de un sector determinado, nos indicaría una afectación del sector topográfico correspondiente del iris.
Así, un vaso grueso y recto indicaría un estado congestivo, un vaso en V indicaría una tendencia espástica y un vaso en U indicaría un edema sanguíneo provocado por un obstáculo.
Existirían una serie de signosgenerales, sin una localización iridológica correspondiente, somo serían los vasos en forma de coma (hipertensión) o zigzagueantes (insuficiencia venosa), pero que, como vemos, ya guardan una relación mucho más estrecha con las esclerología vascular.
Valoraremos los vasos sanguíneos, su disposición, número y estructura.
La esclerología no es un método diagnóstico, sino de valoración pronóstica, valorando la repercusión de la patología sobre todo el sistema vascular.
-EXPLORACIÓN DE LA ESCLERÓTICA
Lo más importante será diferenciar las redes venosas de las arteriales.
Las arteriolas tienen una coloración más pálida y roja que las venas, debido a su menor calibre y a la menor oxigenación de la sangre venosa.
La diferencia de coloración entre arterias y venas es de gran importancia, ya que las venas de color amoratado tienen un especial interés en el diagnóstico.
Lo que nos interesa es el color general de las arterias y venas, indicándonos su grado de oxigenación. Las personas que padecen dificultad de oxigenación de la sangre, suelen tener una tendencia a la coloración oscura de los vasos de la esclerótica.
Las arterias tienen una mayor tendencia a retorcerse, ya que tienen una capa muscular y son de calibre más pequeño que las venas.
La pared de las vénulas es más fina y elástica, estando además desprovista de musculatura, provocando una visión más nítida de su contenido, y en casos anómalos, una mayor facilidad para que el plasma exude hacia el exterior y forme señales de acúmulo como la franja venosa.
El quiasma óptico está localizado muy cercano a otras estructuras dentro del cráneo, tales como la glándula hipófisis, las arterias carótidas internas, los nervios olfatorios y el tercer ventrículo. Existen enfermedades propias del quiasma que pueden alterar de manera especial la visión, pero también algunas enfermedades de las estructuras vecinas que pueden hacerlo. |
|
|
|
La ora serrata es la parte más periférica de la retina, ubicandose en el extremo anterior a la porción central y sensible de la misma que se llama mácula. Esta zona de empalme o confluencia entre la retina y el cuerpo ciliar, conocida como ora serrata, marca la transición entre el área que no es fotosensitiva y la compleja región de varias capas sensibles a la luz.
La zona de la ora serrata es la más periférica de la retina, en contraposición a la porción central y sensible de la misma que se llama mácula .